 我们的研究重点是了解视紫红质和调节其运输的基因突变导致视网膜疾病的分子机制。 视紫红质是视网膜杆状感光细胞感光膜的主要成分,在那里它充当启动光转导级联的感光 GPCR。 视紫红质也是光感受器健康的中央调节剂,并且受到超过 150 种导致视网膜色素变性的不同突变的影响。 影响视紫红质 C 末端的突变会导致严重的常染色体显性视网膜色素变性 (ADRP)。 在我们的研究过程中,我们将视紫质 C 端 VxPx 基序定义为保守的纤毛靶向信号 (CTS)。 我们确定了纤毛靶向复合物的成分,该复合物识别该信号以调节视紫质运输和视网膜杆光感受器膜更新。
我们的研究重点是了解视紫红质和调节其运输的基因突变导致视网膜疾病的分子机制。 视紫红质是视网膜杆状感光细胞感光膜的主要成分,在那里它充当启动光转导级联的感光 GPCR。 视紫红质也是光感受器健康的中央调节剂,并且受到超过 150 种导致视网膜色素变性的不同突变的影响。 影响视紫红质 C 末端的突变会导致严重的常染色体显性视网膜色素变性 (ADRP)。 在我们的研究过程中,我们将视紫质 C 端 VxPx 基序定义为保守的纤毛靶向信号 (CTS)。 我们确定了纤毛靶向复合物的成分,该复合物识别该信号以调节视紫质运输和视网膜杆光感受器膜更新。
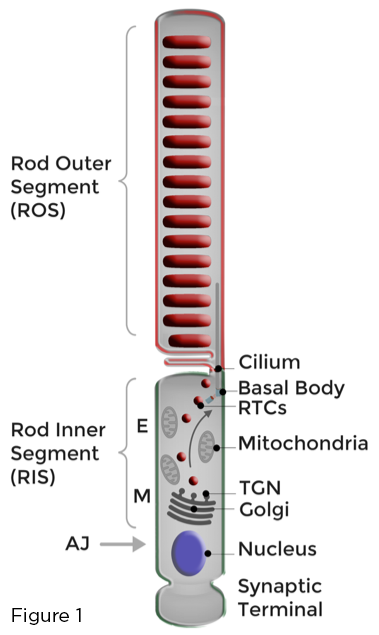 视杆光感受器最佳功能的基础是将载有视紫质的光敏膜严格划分为形成杆外段 (ROS) 的独特修饰的初级纤毛。 通过持续补充光损伤的 ROS 膜和有效隔离参与光转导的 ROS 蛋白和脂质来维持杆状光感受器稳态,远离那些在称为杆内段 (RIS) 的相邻细胞体中发生的各种细胞功能,或在杆状突触末端。 高尔基体和其他生物合成细胞器位于 RIS 中的肌样区域 (M)。 视网膜杆中的极化膜运输涉及合成、分类和运输,通过 RIS,大量高尔基体到纤毛导向的视紫质运输载体 (RTC)。 RTC 穿过充满线粒体的椭圆体区域 (E),并与纤毛附近的 RIS 质膜融合。 然后将新合成的膜传送到 ROS (图 1)。
视杆光感受器最佳功能的基础是将载有视紫质的光敏膜严格划分为形成杆外段 (ROS) 的独特修饰的初级纤毛。 通过持续补充光损伤的 ROS 膜和有效隔离参与光转导的 ROS 蛋白和脂质来维持杆状光感受器稳态,远离那些在称为杆内段 (RIS) 的相邻细胞体中发生的各种细胞功能,或在杆状突触末端。 高尔基体和其他生物合成细胞器位于 RIS 中的肌样区域 (M)。 视网膜杆中的极化膜运输涉及合成、分类和运输,通过 RIS,大量高尔基体到纤毛导向的视紫质运输载体 (RTC)。 RTC 穿过充满线粒体的椭圆体区域 (E),并与纤毛附近的 RIS 质膜融合。 然后将新合成的膜传送到 ROS (图 1)。
分选到 RTC 受 Rab 和 Arf 家族的小 GTP 酶的调节,这些 GTP 酶在组织细胞内膜运输以及向初级纤毛的膜递送中起核心作用。 参与纤毛形成和维持的蛋白质由大约 25% 的遗传性视网膜疾病基因编码,突变会导致视网膜变性、囊性肾、肥胖和神经管缺陷,这些遗传疾病统称为纤毛病。
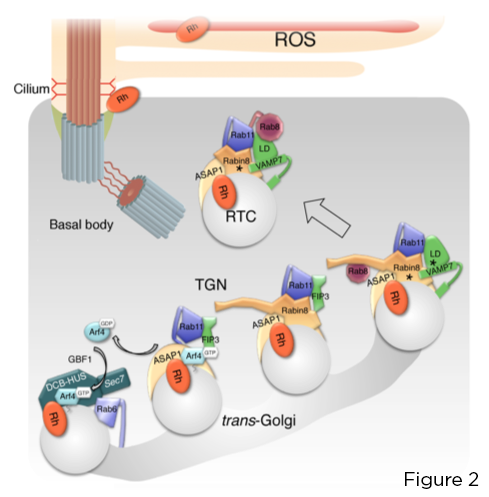 小 GTPase Arf4 识别并直接结合视紫质 C 端 VxPx CTS。 Arf4 由 Arf 鸟嘌呤核苷酸交换因子 (GEF) GBF1 在光感受器高尔基体激活,启动以 Arf GTPase 激活蛋白 (GAP) ASAP1 为中心的靶向关系的逐步组装,后者介导 Arf4 和 Rab11a 上的 GTP 水解-FIP3-Rabin8 双效应复合物。 该复合物控制高度保守的 Rab11a-Rabin8-Rab8 靶向模块的组装,该模块直接将 R-SNARE VAMP7 募集到 RTC 上以通过 VAMP7 与同源质膜 SNAREs 语法蛋白 3 和 SNAP-25 配对来调节它们在纤毛基部的融合(图2)。
小 GTPase Arf4 识别并直接结合视紫质 C 端 VxPx CTS。 Arf4 由 Arf 鸟嘌呤核苷酸交换因子 (GEF) GBF1 在光感受器高尔基体激活,启动以 Arf GTPase 激活蛋白 (GAP) ASAP1 为中心的靶向关系的逐步组装,后者介导 Arf4 和 Rab11a 上的 GTP 水解-FIP3-Rabin8 双效应复合物。 该复合物控制高度保守的 Rab11a-Rabin8-Rab8 靶向模块的组装,该模块直接将 R-SNARE VAMP7 募集到 RTC 上以通过 VAMP7 与同源质膜 SNAREs 语法蛋白 3 和 SNAP-25 配对来调节它们在纤毛基部的融合(图2)。
Rab11a-Rabin8-Rab8 纤毛级联反应的关键是 Rab8 GEF Rabin8,这是一种多功能支架蛋白,可与选择的纤毛蛋白相互作用,例如 TRAPPII 运输复合体和 BBSome,表明在感觉受体的纤毛通路中起核心作用。 其功能受 NDR2 激酶 (STK38L) 突变的影响,该突变由与人类纤毛病 Leber 先天性黑蒙 (LCA) 相对应的犬早期视网膜变性 (erd) 基因编码。 我们目前的研究表明,人类 GFP-Rabin8 在转基因中表达 十条蟾蜍 在高尔基体和 RTC 上与内源性 Rabin8 和视紫质共定位,为未来研究 Rabin8 在沿睫状通路的膜进展中的作用铺平了道路,这可能在遗传性视网膜退行性疾病中受到破坏。
总的来说,我们的研究表明,针对 ROS 的膜靶向是纤毛靶向的一种保守形式。 VxPx 基序存在于其他纤毛膜蛋白中。 保守的基于 Arf4 的靶向复合物通过小 GTP 酶及其调节器的复杂功能网络将感觉受体靶向初级纤毛,这些调节器对导致视网膜变性和纤毛病的突变非常敏感。
Deretic D.、Lorentzen E 和 Fresquez T.(2019 年)。 基于 Arf4 的睫状膜靶向复合物的来龙去脉。 小型 GTP 酶. 小 GTP 酶。 2019 年 9 月 1:12-10.1080。 doi:21541248.2019.1616355/XNUMX。 [Epub 提前印刷]
Kandachar V、Tam BM、Moritz OL 和 Deretic D. (2018) 睫状膜靶向复合物中 SNARE VAMP7 和 Rab-GTPases 之间的相互作用网络。 J Cell Sci. (2018) J 细胞科学。 2018 年 10 月 131 日;24(222034)。 pii:jcs10.1242。 doi:222034/jcs.XNUMX。
Wang J.、Fresquez T. Kandachar V. 和 Deretic D (2017)。 Arf GEF GBF1 和 Arf4 与感觉受体货物视紫质协同调节睫状膜运输。 J Cell Sci. J细胞科学。 2017 年 1 月 130 日;23(3975):3987-10.1242。 doi:205492/jcs.2017。 电子版 12 年 XNUMX 月 XNUMX 日。
Vetter M、Wang J、Lorentzen E 和 Deretic D. (2015) 睫状膜靶向复合物中 Rab11 效应子相互作用网络的新地形。 小 GTP 酶。 2015 年 2 月 6 日;4(165):73-10.1080。 doi:21541248.2015.1091539/2015。 电子版 23 年 XNUMX 月 XNUMX 日。
Wang J 和 Deretic D. (2015) Arf 和 Rab11 效应子 FIP3 与 Arf GAP ASAP1 协同作用以指导 Rabin8 靶向睫状受体。 J Cell Sci. J Cell Sci jcs.162925; 高级在线文章 11 年 2015 月 10.1242 日,doi:162925/jcs.XNUMX (2015)。
Wang J. 和 Deretic D (2014)。 指导视紫质转运至初级纤毛的分子复合物。 Prog Retin Eye Res. 2014 年 38 月;1:19-10.1016。 doi:2013.08.004/j.preteyeres.2013。 电子版 14 年 XNUMX 月 XNUMX 日
Wang J、Morita Y、Mazelova J 和 Deretic D.(2012 年)。 Arf GAP ASAP1 提供了一个平台来调节 Arf4-和 Rab11-Rab8 介导的纤毛受体靶向。 EMBO J 31,4057-4071。
Mazelova J、Ransom N、Astuto-Gribble L、Wilson MC 和 Deretic D. (2009) Syntaxin 3 和 SNAP-25 配对,由 omega-3 二十二碳六烯酸 (DHA) 调节,控制视紫质的递送,用于纤毛的生物发生-衍生感觉细胞器,杆外节。 J.细胞科学。 122,2003-2013。
Mazelova J、Astuto-Gribble L、Inoue H、Tam BM、Schonteich E、Prekeris R、Moritz OL、Randazzo PA 和 Deretic D. (2009) 纤毛靶向基序 VxPx 通过 Arf4 指导贩运模块的组装。 EMBO J. 28,183-192。
Deretic D、Williams AH、Ransom N、Morel V、Hargrave PA 和 Arendt A. (2005) 视紫红质 C 末端,导致视网膜疾病的突变位点,通过与 ARF4 结合来调节贩运。 PROC。 国家科。 科学院。 科学。 美国。 102:3301-3306。